Ilex aquifolium L. (Падуб остролистный, Aquifoliaceae)
 Регистрация даст возможность просматривать полноразмерные фотографии
Регистрация даст возможность просматривать полноразмерные фотографии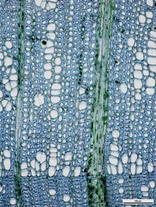 Регистрация даст возможность просматривать полноразмерные фотографии
Регистрация даст возможность просматривать полноразмерные фотографии Регистрация даст возможность просматривать полноразмерные фотографии
Регистрация даст возможность просматривать полноразмерные фотографииПоперечный срез
Древесина рассеяннососудистая. Просветы сосудов не очень многочисленные, собраны в длинные 1–2–3-рядные радиальные цепочки по 2–20 (реже более) клеток, очень редко одиночные. Сосуды ранней древесины могут образовывать прерывистый тангенциальный 1–3-рядный слой. Сосуды мелкие, просветы, в основном слабоугловатые или в разной степени уплощенные.
Переход от ранней древесины к поздней незаметный. К внешней границе кольца прироста просветы сосудов практически не уменьшаются в количестве и по диаметру. Граница кольца прироста более или менее заметная по наличию в конце кольца прироста нескольких рядов сплющенных в радиальном направлении волокнистых трахеид.
Древесинная паренхима скудная апотрахеальная – диффузная, местами соприкасающаяся с группами сосудов.
Лучи узкие, однорядные, практически незаметные на поперечном срезе, и немногочисленные широкие (более широкие или равные группам просветов сосудов).
Основная масса древесины состоит из волокнистых трахеид.
 Регистрация даст возможность просматривать полноразмерные фотографии
Регистрация даст возможность просматривать полноразмерные фотографии Регистрация даст возможность просматривать полноразмерные фотографии
Регистрация даст возможность просматривать полноразмерные фотографииТангенциальный срез
Лучи немногочисленные, однорядные и двух-четырехрядные (до 5). Одно-трехрядные лучи низкие, высотой 1–10 клеток. Многорядные лучи довольно высокие, до 30–40 клеток высотой, иногда с короткими однорядными окончаниями с одной или обеих сторон.
 Регистрация даст возможность просматривать полноразмерные фотографии
Регистрация даст возможность просматривать полноразмерные фотографии Регистрация даст возможность просматривать полноразмерные фотографии
Регистрация даст возможность просматривать полноразмерные фотографии Регистрация даст возможность просматривать полноразмерные фотографии
Регистрация даст возможность просматривать полноразмерные фотографии Регистрация даст возможность просматривать полноразмерные фотографии
Регистрация даст возможность просматривать полноразмерные фотографии Регистрация даст возможность просматривать полноразмерные фотографии
Регистрация даст возможность просматривать полноразмерные фотографииРадиальный срез
Перфорационные пластинки лестничные, с многочисленными (от 15 до 30) перегородками. Межсосудистая поровость очередная, поры многочисленные, мелкие, свободные или сближенные, с округлыми или овальными окаймлениями. По ширине сосуда располагается до 5–7 пор. Сосуды с хорошо заметными спиральными утолщениями. Трахеиды с ясными спиральными или кольцевидными утолщениями и редкими (одно- или, реже, двурядными) окаймленными порами.
Однорядные лучи практически гомогенные, состоят из квадратных и стоячих клеток. Многорядные лучи гетерогенные, из узких лежачих внутренних и крупных квадратных или стоячих краевых клеток
Клетки тяжевой и лучевой паренхимы содержат
многочисленные зерна крахмала.
Ключевые характеристики: Древесина рассеяннососудистая. Просветы сосудов собраны в длинные 1–2–3-рядные радиальные цепочки по 2–20 клеток. Переход от ранней древесины к поздней незаметный. Граница кольца прироста более или менее заметная. Основная масса древесины состоит из волокнистых трахеид. Перфорационные пластинки лестничные, с многочисленными (от 15 до 30) перегородками. Сосуды и трахеиды со спиральными утолщениями. Лучи однорядные и двух–четырехрядные, гетерогенные.
Примечание: Виды рода Ilex по анатомическому строению древесины достоверно не различаются.
 Регистрация даст возможность просматривать полноразмерные фотографии
Регистрация даст возможность просматривать полноразмерные фотографии Регистрация даст возможность просматривать полноразмерные фотографии
Регистрация даст возможность просматривать полноразмерные фотографииОписание: Плод - 4- 5-косточковый пиренарий, образованный верхней завязью 4- 5-членного синкарпного гинецея. Зрелые плоды круглые или продолговатые, 7 - 10 мм длиной и около 8 мм в диаметре, красные или желтые. Косточка 5–7 мм длиной, 3,5–4 мм шириной, 3–3,5 мм толщиной, широкояйцевидная, часто слегка искривленная. Спинка выпуклая, брюшная сторона двугранная с тупым ребром. Поверхность светло-коричневая с желтоватыми разветвленными ребрами, иногда между ними находятся бугорки. (Броувер, Штелин, 2010)
Распространение: эндозоохория (Herrera, 1987, 1989; Debussche, Isenmann, 1992; Williams, Karl, 1996; Obeso, 1998; Arrieta, Suarez, 2005; Martínez et al., 2008; García et al., 2005, 2007, 2016; Møller et al., 2012; Rodríguez -Perez et al., 2014; Ecological...); гидрохоия (Cappers, 1993)
Вес 1000 диаспор, г: 26,5020 (Török ...); 28,7 (García et al., 2007); 40 (Ecological ...; Life form...); 54,1 (Royal...); 140 (Datenbank...) 560,0 плоды, 124, 0 косточки (Williams, Karl, 1996) 26 - 76 косточки (Obeso, 1998); 200 - 300 (Пидотти, 1958); 289,0 плод, 32,2 косточка (Herrera, 1987)
Литература:
Броувер В., Штелин А. Справочник по семеноведению сельскохозяйственных, лесных и декоративных культур с ключом для определения важнейших семян. Пер. с нем. М.: Т-во науч. изд. КМК, 2010. 694 с.
Пидотти О.А. Семена древесных пород // Деревья и кустарники СССР в 6 т. Т. 4. М.; Л.: Изд-во АН СССР. 1958. С. 914 - 917.
Arrieta S., Suarez F. Spatial dynamics of Ilex aquifolium populations seed dispersal and seed bank: understanding the first steps of regeneration // Plant Ecology. 2005. P. 177: 237–248.
Cappers R. T. J. Seed dispersal by water: a contribution to the interpretation of seed assemblages // Vegetation history and archaeobotany. 1993. Vol. 2, № 3. P. 173-186.
Datenbank biologisch-ökologischer Merkmale der Flora von Deutschland. URL:
http://www2.ufz.de/biolflor/overview/gattung.jspDebussche M., Isenmann P. A mediterranean bird disperser assemblage: composition and phenology in relation to fruit availability // Rev. Ecol. (Terre Vie). 1992. Vol. 47. P. 411 – 432.
Ecological flora of British Islands
http://www.ecoflora.co.uk/search_synonyms.phpGarcía D., Carlob T. A., Martínez D. Differential effect of landscape structure on the large-scale dispersal of co-occurring bird-dispersed trees // Basic and applied ecology 2016 v.17 pp. 428-437.
García D., Martínez I., Obeso J. R. Seed transfer among bird-dispersed trees and its consequences for post-dispersal seed fate // Basic and Applied Ecology. 2007. Vol.8. P. 533—543.
García D., Obeso J. R., Martínez I. Blackwell Publishing, Ltd. Spatial concordance between seed rain and seedling establishment in bird-dispersed trees: does scale matter? // J. Ecol. 2005. Vol. 93. P. 693 - 704.
Herrera C.M. Vertebrate-dispersed plants of the Iberian Peninsula: a study of fruit characteristics. // Ecol. Monogr. 1987. Vol. 57. P. 305-331.
Herrera, C.M. (1989): Frugivory and seed dispersal by carnivorous mammals, and associated fruit characteristics, in undisturbed Mediterranean habitats. Oikos 55: 250-262.
Life form, breeding system and seed mass for 1116
Martínez I., García D., Obeso J. R. Differential seed dispersal patterns generated by a common assemblage of vertebrate frugivores in three fleshy-fruited trees // Ecoscience 2008. Vol. 15, N 2. P. 189-199.
Møller L. A., Skoua A.-M. T., Kollmann J. Dispersal limitation at the expanding range margin of an evergreen tree in urban habitats? // Urban Forestry & Urban Greening. 2012. Vol. 11. P. 59–64.
Obeso J. R. (1998) Patterns of variation in Ilex aquifolium fruit traits related to fruit consumption by birds and seed predation by rodents, Écoscience, 5:4, 463-469.
Rodríguez -Perez J., García D., Martínez D. Spatial networks of fleshy-fruited trees drive the flow of avian seed dispersal through a landscape // Functional Ecology. 2014. Vol. 28. P. 990–998.
Royal Botanic Gardens Kew. (2008) Seed Information Database (SID). Version 7.1. Available from: http://data.kew.org/sid / (May 2008)
Török P., Miglécz T., Valkó O., Tóth K., Kelemen A., Albert Á.-J., Matus G., Molnár A.V., Ruprecht E., Papp L., Deák B., Horváth O., Takács A., Hüse B., Tóthmérész B. New thousand-seed weight records of the pannonian flora and their application in analysing social behaviour types // Acta Bot. Hungarica. 2013. Vol. 55, № 3–4. P. 429–472.
Williams P. A., Karl B. J. Fleshy fruits of indigenous and adventive plants in the diet of birds in forest remnants, Nelson, New Zealand // New Zealand Journal of Ecology (1996) 20(2): 127-145.
Этикетка: Батуми, БС, 1950